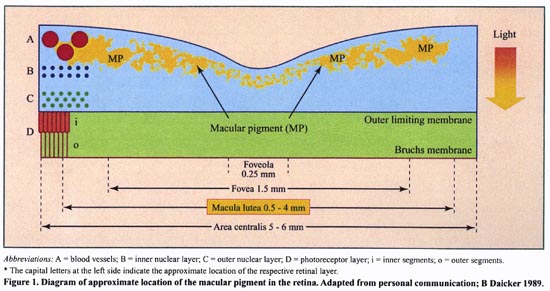
 Schéma Macula lutea
Schéma Macula lutea
La dégénérescence maculaire liée à l’âge (DMLA) est la cause la plus fréquente de mal-voyance et de cécité, touchant les personnes de plus de 60 ans, dans les pays industrialisés.
En France, on estime que plus d’un million de personnes serait touché par la DMLA (syndicat national des ophtalmologistes français). Ce nombre est en constante augmentation, ne serait-ce que par l’allongement de la durée de la vie. Il serait même multiplié par 3 d’ici à 25/30 ans. Globalement, + de 15% de la population occidentale serait touchée par cette pathologie.
Cette maladie, sous sa forme sèche notamment, a une réputation d’irréversibilité, sans traitement vraiment retenu à ce jour.
La prévention et les possibilités de correction de la DMLA apparaissent de ce fait d’un intérêt majeur pour la santé.
Rappelons-en les grandes lignes :
- La rétine est riche en acides gras essentiels polyinsaturés (AGPI) oméga 3 (EPA/DHA).
- Elle est donc potentiellement et particulièrement exposée aux attaques radicalaires oxydatives.
- La fovéa appelée aussi macula lutea concentre naturellement 2 caroténoïdes : Lutéine et Zéaxanthine. Cette concentration est unique.
- Les différentes études (Siems WG) nous montrent que l’oxydation des caroténoïdes au soleil et lumière UV nous donne une échelle de dégradation allant de Lycopène> Zéaxanthine > Lutéine
- Ceci nous confirme que la Lutéine est spécifiquement la plus résistante face aux attaques radicalaires UV. Cela explique la « relative absence » du Lycopène dans la macula alors qu’il est le caroténoïde le plus concentré dans le sérum. Et ce, parallèlement à une très forte concentration de Lutéine dans la fovéa (2,7 fois plus), déclinant très rapidement à la périphérie.
- On constate aussi que la Lutéine est en plus forte concentration naturellement, avec macula saine, dans les « rétines jeunes » / « rétines âgées ».
- Il apparaît (Sujak et al.) aussi que la Zéaxanthine est d’avantage impliquée dans la protection de la lipoperoxydation et la Lutéine dans la protection liée aux radiations photoniques à onde courte pénétrant dans la rétine.
- De même, on constate une inégalité de risque entre les hommes et les femmes : les hommes auraient une densité pigmentaire de +38% / femmes et, ceci en dépit d’une concentration plasmatique semblable. Le facteur diététique notamment lié aux graisses, pourrait être incriminé ainsi, que les habitudes tabagiques liées.
Toutes ces données nous confortent dans l’effet protecteur, maintenant clairement identifié par les chercheurs, de la Lutéine dans la prévention de la DMLA.
Parallèlement à ces études sur la réalité préventive de la Lutéine, un certain nombre de chercheurs (Handelman GJ, Dratz EA, Reay CC, van Kuijk JG,..) a élaboré ces dernières années l’hypothèse selon laquelle, la synergie d’action d’antioxydants pourrait dans certains cas aller au delà du stade préventif pour donner un début de réponse correctrice dans les DMLA.
Reprenant en cela certaines études (non certifiées à l’époque) et surtout les exposés de cas dans lesquels il apparaissait que cette synergie d’action modifiait le décours de cette maladie.
Quelques études récentes, certifiées, nous apportent un certain nombre de réponses sur ce qu’envisageaient les hypothèses précédentes : Savoir qu’une supplémentation en Lutéine en synergie ou non avec d’autres antioxydants, notamment des caroténoïdes, pouvait non seulement différer le risque mais, aussi détourner, modifier le cours d’une DMLA acquise.
Apports suggérés par une supplémentation chez des sujets sains et atteints de dégénérescence oculaire (cataracte/DMLA).
a. Chez le sujet sain :
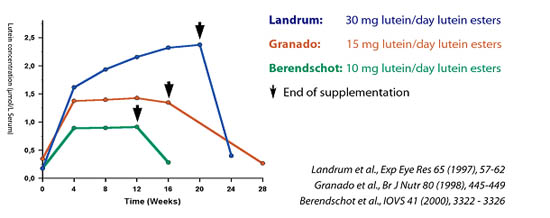 Schéma Landrum, Granado, Berendschot
Schéma Landrum, Granado, BerendschotLandrum, Granado et Berendschot ont supplémenté des patients sains pendant plusieurs mois aux doses respectives de 30, 15 et 10 mg Lutéine / jour. Ces trois doses rechargent rapidement (4 semaines) les taux sanguins et durablement la densité optique du pigment maculaire.
Ces études d’intervention (supplémentation en doses > 6 mg nutritionnelles optimales) ont été réalisées sur des périodes de 12 à 36 semaines, les apports totaux cumulés en Lutéine libre varient de ± 720 mg à ± 4320 mg.
Landrum a donné 4200 mg de Lutéine libre (8400 mg d’esters) au cours des 140 jours de supplémentation, la teneur en Lutéine s’est élevée de 10x entre les 20ème et 40ème jours de supplémentation. Après cette rapide augmentation de la teneur sanguine, celle-ci s’est stabilisée en plateau. La densité optique du pigment maculaire s’est élevée à partir de 20ème au 40ème jour et a même continué à augmenter durant 40 à 50 jours après l’arrêt de la supplémentation.
Granado et Berendschot ont montré qu’une supplémentation par 15 mg/jour et 10 mg/jour élevait d’un facteur 5 le taux sérique en Lutéine . Selon Granado, une supplémentation continue par 15 mg de Lutéine /jour induit un « ceiling effect » qui traduit la saturation de la capacité de transport de la Lutéine par les lipoprotéines. Cette saturation semble atteinte pour une concentration sérique > 1,05 µmol/l.
La Lutéine se ré-estérifie et se stocke dans les lipoprotéines.
Berendschot estime que la dose de 10 mg/jour est suffisante pour élever d’un facteur 5 le taux sérique et de 4 à 5% la densité optique du pigment maculaire.
D’une manière générale, ces études montrent que le pigment maculaire continue à s’élever pendant quelques semaines après la supplémentation, ce qui explique la constitution d’une mise en réserve en Lutéine et d’un relatif faible turn-over des xanthophylles dans les pigments rétiniens en post-réplétion.
b. Chez le sujet atteint de dégénérescence oculaire :
Les études les plus récentes nous informent que chez les patients atteints de dégénérescence oculaire, Olmedilla a donné 4320 mg de Lutéine libre à raison de 45 mg par semaine (15 mg tous les 3 jours, soit ± 6,5 mg/jour) ; ce qui équivaut à 3 repas riches en Lutéine par semaine : la teneur sérique a progressé pendant 12 mois et s’est stabilisée au cours de la 2ème année de supplémentation.
Olmedilla B.et al. ont montré aussi qu’une supplémentation par 45 mg/semaine élève de façon linéaire la concentration sanguine en Lutéine et a permis l’observation d’améliorations cliniques intéressantes, à savoir : l’augmentation de l’acuité visuelle de ± 64% et l’augmentation de la sensibilité à la lumière de 33%.
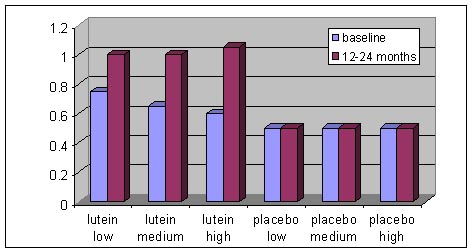 Schéma Lutein low
Schéma Lutein lowS. Richer et col. ont démontré l’intérêt d’une dose de 10mg de Lutéine (libre) administrée seule ou en association avec des antioxydants chez des patients atteints de DMLA atrophique.
Les caroténoïdes et, particulièrement la Lutéine , apparaissent ainsi en supplémentatation synergique ou non, capables de modifier, détourner le cours de cette maladie. Face à ce véritable problème de santé publique que représente la DMLA, Zéaxanthine et Lutéine représentent, à ce jour, une des rares réponses positives certifiées.
Références bibliographiques sur demande à Cette adresse e-mail est protégée contre les robots spammeurs. Vous devez activer le JavaScript pour la visualiser.
c. La lutéine et l'ischémie vasculaire :
Les patients diabétiques, ou souffrant du syndrôme métabolique présentent des risques élevés de développer des pathologies cardiovasculaires liées à l’hyperlipidémie et corrélées à un état pro-inflammatoire.
Des publications récentes ont mis en évidence le rôle des micro-nutriments cellulaires (Lutéine, DHA, α-tocophérol, Sélénium, Zinc,…) dans la régulation métabolique et physiologique du système cardiovasculaire.
J. Dwyer et al. ont montré que la Lutéine était très active dans la réduction des LDL oxydées et dans l’inhibition de la réponse inflammatoire des monocytes aux LDL piégées dans la paroi artérielle.
Chez des souris apo E-null et LDL-receptor-null, une supplémentation en Lutéine a réduit les hydroperoxydes lipidiques plasmatiques de 78%, et la taille des lésions aortiques de 44% par rapport au contrôle. Ces auteurs ont également objectivé l’inhibition in vitro de la chémo-attraction des monocytes par l’addition de Lutéine dans un milieu de culture de cellules endothéliales humaines. The Los Angeles Atherosclerosis Study. Circulation 2001.
Cette propriété protectrice de la Lutéine a été évaluée dans une étude prospective réalisée sur les 480 sujets de la cohorte de la Los Angeles Atherosclerosis Study; ces participants n’avaient aucun antécédent d’infarctus du myocarde, d’angor ou d’AVC. Dwyer et ses collaborateurs ont mesuré chez ces sujets, par ultrasono-graphie, l’épaisseur de l’intima-media carotidien (IMT) durant 18 mois et dosé les teneurs plasmatiques en Lutéine (L) et zéaxanthine (Z). Les sujets ayant les quintiles les plus élevés en (L) et (Z) ont développé une progression de l’IMT quasi nulle (0,004 mm) alors que les individus avec le quintile inférieur ont progressé de 0,021 mm (p=0,01).The Los Angeles Atherosclerosis Study. Arterosclerosis, Thrombosis and Vascular Biology. 2004
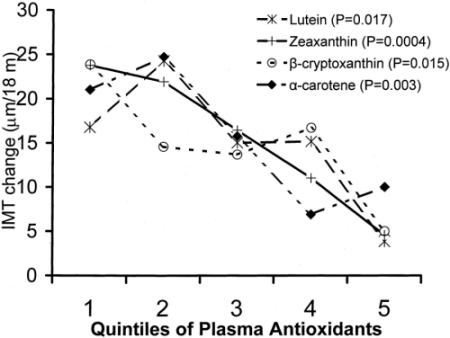 Schéma of Plasma carotenoid quintile
Schéma of Plasma carotenoid quintile
Plasma carotenoid quintiles by 18-month IMT change (adjusted for age, sex, antioxidant and sex interaction, and smoking status). The Los Angeles Atherosclerosis Study. Probability values are for trend. Antioxidant quintile ranges (µmol/L): lutein: 0.02 to 0.18, 0.18 to 0.23, 0.24 to 0.29, 0.29 to 0.36, 0.36 to 0.81; zeaxanthin: 0.01 to 0.04, 0.04 to 0.05, 0.05 to 0.06, 0.06 to 0.08, 0.09 to 0.28; ß-cryptoxanthin: 0.01 to 0.04, 0.05 to 0.06, 0.06 to 0.08, 0.08 to 0.12, 0.12 to 0.60; -carotene: 0.01 to 0.07, 0.07 to 0.11, 0.11 to 0.15, 0.16 to 0.27, 0.27,1.06; ß-carotene: 0.02 to 0.24, 0.24 to 0.42, 0.42 to 0.64, 0.64 to 1.04, 1.05 to 8.00; and lycopene: 0.03 to 0.31, 0.31 to 0.47, 0.47 to 0.67, 0.67 to 1.02, 1.03 to 6.47.
____________
Les observations de Dwyer confirment celles de C. Iribaren et al qui avaient montré la corrélation inverse entre les teneurs plasmatiques en Lutéine et zéaxanthine et l’épaississement précoce de l’intima media des carotides. The ARIC Study. Arteriosclerosis, Thrombosis and Vascular Biology 1997.
Lors des infarctus du myocarde et des accidents vasculaires cérébraux, de nombreux radicaux libres issus du métabolisme de l’oxygène et de l’azote sont libérés, ils constituent des facteurs de risques importants et souvent décisifs dans le pronostic et la survie du patient. A. Cherubini et C. Polidori ont dosé les teneurs plasmatiques de caroténoïdes dont la Lutéine et la zéaxanthine et le MDA (Malonedialdéhyde = le marqueur de la peroxydation des lipides membranaires) chez 28 patients, lors de leur admission après un AVC. Ces patients ont été répartis en deux groupes : un groupe « stable » (sans déclin fonctionnel) et un groupe « aggravé » selon le Barthel index. Les teneurs en caroténoïdes ont chuté au cours des premières 24 heures suivant l’admission, et sont remontées par la suite vers le 7ème jour.
Dans le groupe « stable », les teneurs en Lutéine étaient plus élevées que dans le groupe « aggravé », de plus, les teneurs en MDA étaient significativement plus faibles dans le groupe «stable » par rapport au groupe « aggravé ». Selon ces auteurs, la Lutéine est significativement corrélée à l’issue des patients après AVC et à la sévérité de l’AVC.
C. Polidori et al. ont observé la même corrélation Lutéine et MDA chez 30 patients atteints d’infarctus du myocarde. Une étude clinique suédoise a déterminé les taux plasmatiques de 6 caroténoïdes chez 39 patients atteints de maladie coronarienne aiguë, chez 50 patients avec maladie coronarienne stable et 50 sujets contrôles. Les deux groupes présentaient des taux plasmatiques de Lutéine et zéaxanthine significativement inférieurs à ceux du groupe témoin. Ces teneurs basses en Lutéine étaient associées au tabac, à un BMI élevé, de faibles teneurs en HDL et à un état inflammatoire. Les valeurs plasmatiques de Lutéine étaient de façon indépendante associées aux teneurs en natural killer (NK) dans le sang. Cette relation entre les NK et les oxycaroténoïdes indique le rôle particulier joué par la Lutéine dans le processus immunologique des maladies coronariennes.
Les études épidémiologiques montrent l’effet protecteur des hydroxy-caroténoïdes (Lutéine, Zéaxanthine, Cryptoxanthine) vis-à-vis des maladies cardio-vasculaires.
Howard et al, ayant observé une incidence plus faible des pathologies cardiovasculaires chez les Toulousains que chez les habitants de Belfast, ont comparé les teneurs plasmatiques en Lutéine de ces deux populations ; celles des Toulousains étaient deux fois plus élevées (0.54 μmole/L) ce qui constitue environ un apport alimentaire quotidien de 3mg de Lutéine. (300 g légumes/jour et 250 g fruits/jour). Etude MONICA Toulouse-Belfast. Internat.J.Vit.Nutr.Res. 1996.
Dans une étude prospective menée pendant 7,2 ans sur 638 sujets âgés de 65-85 ans, F.G. De Waart et al. concluent que les teneurs sériques en Lutéine, zéaxanthine et ß-cryptoxanthine sont inversément associées à toutes les causes de mortalité observées dont les maladies cardio-vasculaires et les cancers.
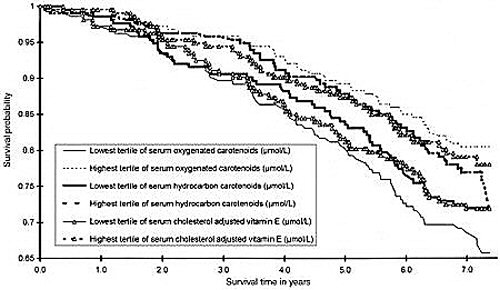 Schéma of Kaplan-Meier estimates of survival functions
Schéma of Kaplan-Meier estimates of survival functions
Kaplan-Meier estimates of survival functions for all-cause mortality of the lowest and highest tertiles of serum oxygenated and hydrocarbon carotenoids and cholesterol adjusted serum vitamin E
Références bibliographiques sur demande à Cette adresse e-mail est protégée contre les robots spammeurs. Vous devez activer le JavaScript pour la visualiser.
d. La lutéine et les pathologies vasculaires associées au diabète et au vieillissement cellulaire :
De nombreuses études ont établi le lien entre les pathologies rétiniennes diabétiques, la cataracte, la dégénérescence maculaire liée à l’âge (DMLA), les maladies cardio-vasculaires et la Lutéine.
Au Japon, the Funagata Study, réalisée sur 1638 sujets souffrant de rétinopathie avec des altérations micro-vasculaires associe le syndrome métabolique identifié chez certains patients, aux altérations micro-vasculaires de la rétine. The Funagata Study : The metabolic syndrome and retinal microvascular signs in a Japan population. B.J. Ophtalmology 2008
Une étude australienne a suivi 2335 sujets pendant 5 ans et 1952 sujets pendant 10 ans ; les patients atteints de DMLA précoce avaient un risque ratio (RR) de 2.32 soit une prédiction de doublement de la mortalité cardiovasculaire. La DMLA tardive prédisait un RR de 5.57 pour la mortalité cardiovasculaire et un RR de 10.21 pour la mortalité par AVC. Cette étude a montré que la DMLA était un facteur prédictif des AVC et pathologies cardiovasculaires sur le long terme chez des patients de 49 à 75 ans.
Dans une autre étude australienne, réalisée sur 1021 sujets d’âge moyen ayant un diabète de type 2, mais sans trouble de la fonction rénale et sans pathologie coronarienne, 125 sujets avaient une rétinopathie et 106 (10.1 %) ont eu un infarctus du myocarde au cours des 9 années qui ont suivi. Selon ces auteurs, la présence d’une rétinopathie diabétique signifie un risque important d’infarctus, indépendant des autres facteurs de risque connus.
D’autres études, dont l’Age-Related Mucular Degeneration : Risk Factors Study Group. (Arch. Opthalmol.2000) et The Rotterdam Study (Am.J. Epidemiol.1995) ont suggéré une étiologie commune à l’athérosclérose responsable des AVC, maladies cardiovasculaires et à la néovascularisation de certaines formes de DMLA.
Dans l’étude CARDIA, réalisée sur 4493 sujets pendant 15 ans, des concentrations élevées en caroténoïdes sériques sont associées à un faible risque de diabète de type 2 et de résistance à l’insuline chez les non-fumeurs exclusivement. The Coronary Artery Risk Development in young adults (CARDIA) Study. American Journal of Epidemiology 2006.
R. Kowluru et al. suggèrent qu’une supplémentation en zéaxanthine chez les patients diabétiques peut être utile dans la prévention ou retarder le développement des rétinopathies. Les supplémentations en zéaxanthine chez le rat ont, par l’action anti-oxydante, inhibé la peroxydation des phospholipides membranaires des photorécepteurs et inhibé l’histopathologie capillaire caractéristique des rétinopathies diabétiques.
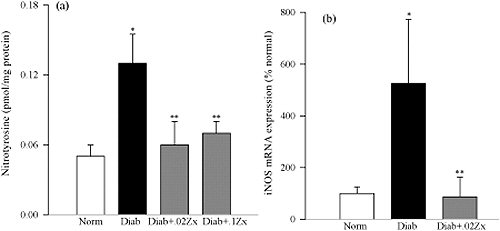 Schéma Effect of Zx on retinal nitrotyrosine and iNOS expression.
Schéma Effect of Zx on retinal nitrotyrosine and iNOS expression.Effect of Zx on retinal nitrotyrosine and iNOS expression. (a) Nitrotyrosine concentrations in the retina were quantified with a nitrotyrosine EIA kit. Each sample was measured in duplicate. The data are expressed as the mean ± SD of results in six rats each in the normal and diabetes+Zx groups and seven rats in diabetes group. (b) mRNA content of iNOS was measured using Q-RT-PCR and gene expression for iNOS was determined in the retina. The levels of iNOS mRNA were normalized to that of the housekeeping gene, B2M, in the same sample. The levels measured in normal rat retinas were considered to be 100%. Data are expressed as the mean ± SD of results in five or more rats in normal, diabetes, and diabetes+0.02% Zx. *P < 0.05 compared with the normal group; **P < 0.05 compared with the diabetes group.
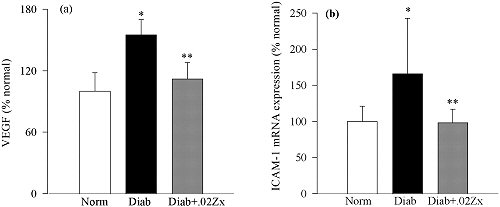 Schéma Effect of Zx on the diabetes-induced increase in retinal growth factors
Schéma Effect of Zx on the diabetes-induced increase in retinal growth factors
Par ailleurs, les propriétés anti-inflammatoires in vitro de la Lutéine s’expliquent par une diminution de l’expression des iNOS (inductible nitric oxide synthase). Dans des cellules de macrophages de souris RAW 264.7, la Lutéine diminue de 50% la production de NO induite par les lipopolysaccharides (LPS) ; l’expression au niveau du mRNA de l’iNOS est réduit de 1.9 fois par la Lutéine, au niveau protéique, l’expression de iNOS induite par le LPS est diminuée de 72.5 % par la Lutéine.
La Lutéine inhibe l’expression de l’iNOS , de la COX2, elle empêche l’activation du NF-kappa B et régule ainsi l’expression des protéines et cytokines pro-inflammatoires inductrices d’angiogénèse et de néovascularisation observées chez les sujets diabétiques, et dans certaines formes de DMLA.
Xue-Hai Jin et al ont mis en évidence cette action anti-inflammatoire de la Lutéine dans un modèle d’uvéite induite par endotoxines (LPS).
Ces propriétés anti-inflammatoires ont été confirmées par J.H. Kim et al qui ont montré l’inhibition par la Lutéine de l’expression des gènes du NF-kappa ß au travers d’une régulation Redox du phosphatidylinositol 3-kinase (PTEN/Akt) et de la voie des kinases inductrices de la libération du NF-kappa B. La translocation du NF-kappa B sur le noyau étant ainsi bloquée, la synthèse des protéines pro-inflammatoires et des molécules d’adhésion (ICAM, VCAM,…) est rendue non fonctionnelle.
La Lutéine réduit in vivo l’accumulation intracellulaire des métabolites pro-oxydants par destruction de l’anion superoxyde O2.- et du peroxyde d'hydrogène l’H2O2.
Les ischémies, le stress oxydant produisent des altérations neuronales irréversibles dans les pathologies oculaires telles que le glaucome et les rétinopathies diabétiques.
Au cours des ischémies, on observe une déplétion en ATP, une excitotoxicité par le glutamate, qui conduisent à la mort des RGC (rétinal ganglion cells). La reperfusion qui suit l’ischémie provoque un stress oxydant par la libération de radicaux libres responsables des altérations neuronales (RGC). Suk-Yee Li and Amy C.Y. Lo ont démontré l’effet protecteur in vivo de la Lutéine, vis-à-vis des effets néfastes de l’ischémie-reperfusion. Cet effet neuroprotecteur de la Lutéine a été objectivé par une hypoxie induite par le CoCl2 et par l’ H2O2 sur des RGC.
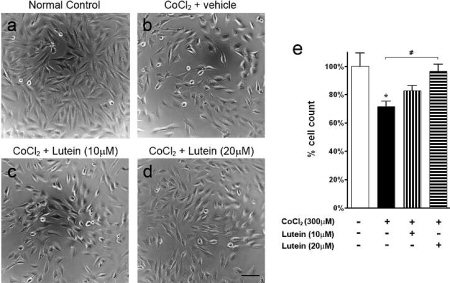 Schéma Light micrographs and cell count of RGC-5 cells treated with cobalt chloride
Schéma Light micrographs and cell count of RGC-5 cells treated with cobalt chlorideLight micrographs and cell count of RGC-5 cells treated with cobalt (II) chloride (CoCl2; 300 μM). (a) Normal control. (b) Vehicle treatment. (c) Lutein treatment at 10 μM. (d) Lutein treatment at 20 μM. CoCl2-induced hypoxia led to cell death in the vehicle-treated group (b) compared with control (a). However, 20 μM lutein treatment reversed the cytotoxic effect of CoCl2 (d). (e) Count of RGC-5 cells treated with CoCl2 referenced to the normal control. A decreased cell number was observed for the vehicle-treated group (*p < 0.05 versus control). However, an increased RGC-5 cell number was observed after 20μM lutein treatment (#p < 0.05 versus vehicle-treated). Scale bar, 25 μm. Error bars, SEM.
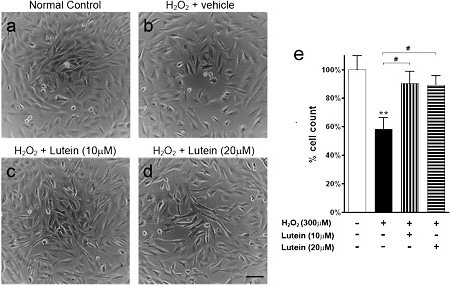 Schéma Light micrographs and cell count of RGC-5 cells treated with hydrogen peroxide
Schéma Light micrographs and cell count of RGC-5 cells treated with hydrogen peroxide
Dans un modèle d’ischémie-reperfusion de la rétine, Suk-Yee Li et al viennent dans une étude récente d’élucider l’effet neuroprotecteur de la Lutéine par son inhibition de l’expression de la nitrotyrosine (NT) et du poly (ADP-ribose) (PAR). La NT est un marqueur de l’oxydation neuronale, elle résulte de la réaction d’oxydation du peroxynitrite sur des résidus de Tyrosine protéique. L’hyperproduction de peroxynitrite active la PAR-polymérase qui libère le PAR, un facteur nucléaire d’immunoréactivité et inducteur d’aptose neuronale.
L’action de la Lutéine au sein des cellules ganglionnaires neuronales se focalise en particulier au niveau de la tête du nerf optique (au centre de la rétine). Cette action antioxydante spécifique dans ce tissu oculaire est due à la structure spatiale d’oxycaroténoïdes de la Lutéine dont les deux radicaux hydroxyles autorisent un « ancrage » dans les phospholipides membranaires du tissu rétinien. Cette particularité « spatiale » de la Lutéine fait de cet oxycaroténoïde un anti-oxydant « tissu spécifique » plus efficace contre les radicaux libres (anion superoxyde, H2O2, peroxynitrite, …) que les autres caroténoïdes non-hydroxylés (ß-carotène, lycopène,…)
Cet effet neuroprotecteur de la Lutéine a également été confirmé par les travaux de M. Sasaki et al. Selon ces travaux, la Lutéine prévient la diminution de l’expression de la Rhodopsine, de GFAP (glial fibrillary acid protein des cellules gliales Müller) induite par endotoxine (LPS).
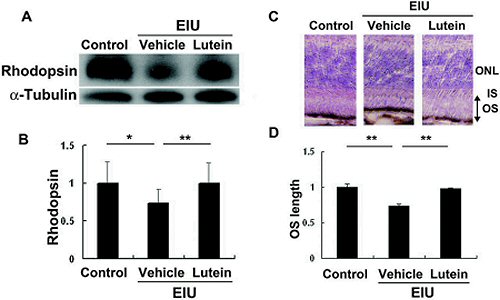 Schéma Inhibitory effect of lutein on EIU-induced decrease in rhodopsin protein and OS length
Schéma Inhibitory effect of lutein on EIU-induced decrease in rhodopsin protein and OS length
Une étude récente menée en Australie sur 111 sujets diabétiques type 2, estime que les teneurs élevées en Lutéine / zéaxanthine et lycopène sont prédictives de faibles risques de rétinopathie diabétiques après ajustement des facteurs confondants et des facteurs de risque. Les concentrations sériques de Lutéine/zéaxanthine + lycopène étaient significativement plus faibles dans le groupe de patients diabétiques avec rétinopathie (OR 1.2).
e. La lutéine et le tissu adipeux pro-inflammatoire :
Le mécanisme par lequel l’obésité est reliée à la DMLA, aux rétinopathies, à la cataracte, s’explique par une modification du métabolisme de la Lutéine et du profil des lipoprotéines véhiculant la lutéine dans le tissu adipeux et dans les tissus oculaires.
Références bibliographiques sur demande à Cette adresse e-mail est protégée contre les robots spammeurs. Vous devez activer le JavaScript pour la visualiser.
ATHEROSCLEROSE, ALZHEIMER, DMLA, SYNDROME METABOLIQUE : ANALOGIES ETIOLOGIQUES ET POLYMORPHISME GENETIQUE. TROPISME SPECIFIQUE DE LA LUTEINE
L’athérosclérose, la maladie d’Alzheimer, la DMLA, le syndrome métabolique sont des maladies caractérisées par un état inflammatoire chronique associé le plus souvent au trouble du métabolisme lipidique. Ces pathologies dégénératives sont induites par un déséquilibre en antioxydants physiologiques.
Les polymorphismes des gènes régulant le métabolisme lipidique sont également des facteurs impliqués dans l’étiologie de ces maladies dégénératives.
La Lutéine, le sélénium, le zinc, l’α-tocophérol sont des nutriments essentiels au maintien du statut antioxydant au sein des cellules spécifiques concernées par cette dégénérescence fonctionnelle ; l’α-tocophérol et la Lutéine sont particulièrement impliqués dans la protection anti-radicalaire des lipoprotéines HDL, LDL et VLDL. De plus, la Lutéine a un tropisme spécifique pour les lipoprotéines plasmatiques et les lipides de l’épithélium pigmentaire rétinien.
a. Analogies de la physiopathologie entre athérosclérose et DMLA :
Les mécanismes impliqués dans la genèse des lésions athéromateuses présentent des similitudes physiopathologiques entre la DMLA et le processus athéromateux.
Dans ces deux pathologies dégénératives, le rôle pro-athérogène du cholestérol et des oxy-stérols a été largement démontré par leurs activités cytotoxiques, pro-oxydantes et pro-inflammatoires.
Ces similitudes physiopathologiques se retrouvent également dans les maladies neurodégénératives dont Alzheimer.
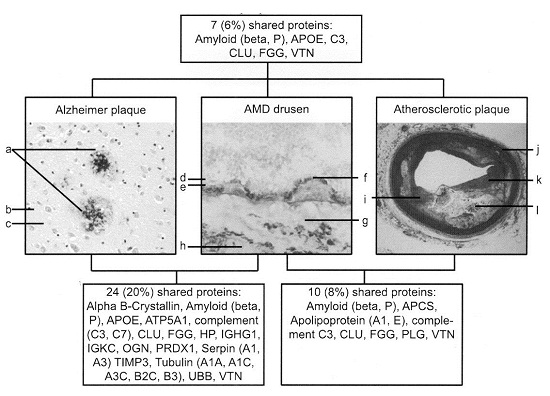 Overlap in protein content of Alzheimer plaques, AMD drusen, and atherosclerotic plaques.
Overlap in protein content of Alzheimer plaques, AMD drusen, and atherosclerotic plaques.
a.1. Analogies histologiques et anatomopathologiques :
On observe dans ces deux pathologies, un dysfonctionnement des cellules endothéliales vasculaires, accompagné de néovascularisation, d’angiogénèse. Dans l’athérosclérose, les dépôts lipidiques dans l’intima des parois vasculaires sont l’élément initial qui induit ces lésions fibro-lipidiques. Celles-ci produisent un épaississement avec perte d’élasticité des parois vasculaires par dénaturation des fibres de collagène.
Par analogie avec l’athérosclérose, les drusen se forment au contact de la membrane de Bruch composée de couches fibro-élastiques de collagène et des cellules endothéliales des capillaires choroïdiens. L’accumulation des dépôts lipidiques renforce le caractère hydrophobe de la membrane de Bruch, ce qui réduit les échanges entre l’épithélium pigmentaire rétinien et la choroïde.
a.2. Analogie du trouble du métabolisme lipidique :
Dans l’athérogénèse, les cellules endothéliales interagissent avec les lipo-protéines plasmatiques. Des cellules endothéliales régulent le transport trans-endothélial des lipoprotéines. Lorsque ces cellules sont intactes, elles évitent une accumulation intimale de dépôt de cholestérol.
Lorsque la perméabilité des cellules est altérée notamment par un état pro-inflammatoire généré par un excès de radicaux libres oxygénés ou nitrés, l’équilibre entre influx et efflux de LDL est rompu. Les LDL, les VLDL en particulier lors de leur entrée dans l’espace sous – endothélial, produisent des LDL oxydées qui seront avec les macrophages, les cellules endothéliales, les cellules musculaires lisses, les différents acteurs de l’athérosclérose.
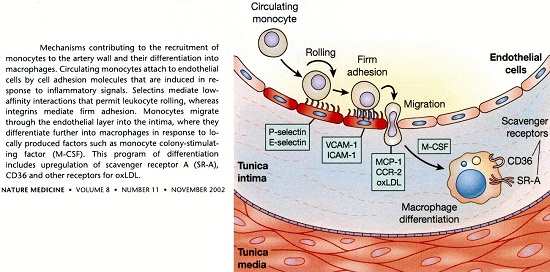 Mechanism contributing to the recruitment of monocytes
Mechanism contributing to the recruitment of monocytes
De même, dans la DMLA, l’épithélium pigmentaire rétinien subit l’action pro-oxydante et pro-inflammtoire des radicaux libres lorsqu’ils sont produits en excès lors de déplétion des anti-oxydants physiologiques (GSH, SOD, vitamine C, tocophérols, lutéine, zinc, sélénium).
L’épithélium pigmentaire comme les cellules endothéliales vasculaires est particulièrement exposé à un environnement riche en oxygène, il est de plus soumis à une exposition prolongée aux rayons lumineux générateurs d’oxygène singulet et à la phagocytose peroxydative (H2O2) des macrophages assurant la maintenance quotidienne des photorécepteurs.
Dans la couche de collagène interne de la membrane de Bruch au contact des cellules de l’épithélium pigmentaire se forment des dépôts de lipides de glycoprotéines et de protéines. Les dépôts focaux constituent les drusen, dont les drusen séreux seront à la base des formes néovasculaires de la DMLA.
Le rôle pro-inflammatoire des drusen présents dans la DMLA peut être assimilé à celui des dépôts lipidiques de l’intima des plaques d’athérome.
Dans l’étiogénèse de ces deux pathologies dégénératives, le rôle délétère de certains oxystérols a été mis en évidence.
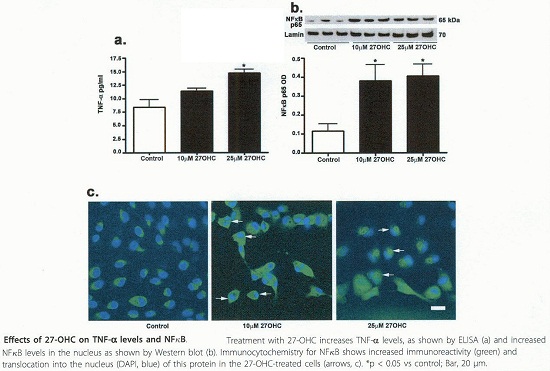 Effects of 27-OHC on TNF levels and NFKB
Effects of 27-OHC on TNF levels and NFKB
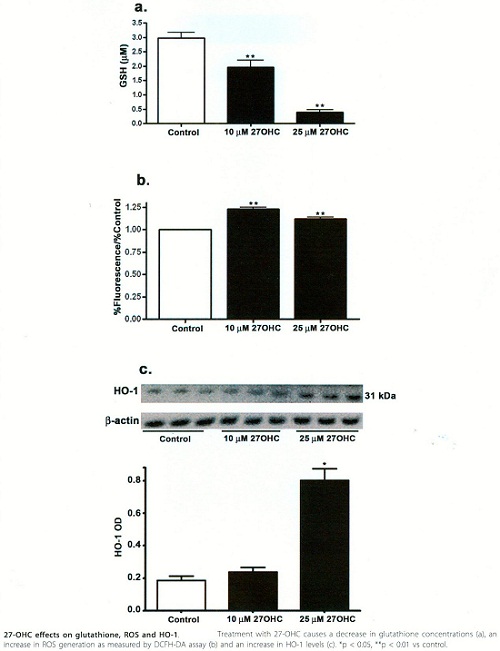 27 - OHC effects on glutathione, ROS and HO-1
27 - OHC effects on glutathione, ROS and HO-1
Les oxystérols sont des dérivés du cholestérol dont les 7-cétocholestérol, 7β-hydroxystérols et 27-hydroxystérols sont cytotoxiques, pro-oxydants et pro-inflammatoires. Les 17-céto et 7β-OH induisent une mort par apoptose des cellules endothéliales des cellules musculaires lisses et des macrophages.
Ces oxystérols sont également impliqués dans la surproduction de radicaux libres, d’IL8 dans les monocytes et dans les cellules des muscles lisses.
Par analogie, les oxystérols produits dans les dépôts lipidiques de la membrane de Bruch par internalisation des LDL oxydés exerceraient ainsi les mêmes effets délétères que dans les parois vasculaires.
Des études récentes ont mis en évidence le rôle des oxystérols dans la DMLA.
Les cellules de l’épithélium pigmentaire rétinien (RPE) expriment des récepteurs aux lipoprotéines oxydées autorisant leur internalisation. L’APO B100 a également été identifiée dans les drusen de patients atteints de DMLA.
![]() Lipid transport and 7 KCh metabolism in the retina
Lipid transport and 7 KCh metabolism in the retina
Le mRNA de l’APO B100 et l’APO B100 ont aussi été mis en évidence dans le RPE, ce qui indique une capacité de synthèse des LDL par ces cellules, l’APOE est présente dans la rétine, elle y est synthétisée dans les cellules de Müller et dans l’épithélium pigmentaire.
L’internalisation des LDLox produit une cytotoxicité médiée par le 7-céto-oxystérol.
De même, le récepteur OSBP (Oxysterol-Binding-Protein) présente une forte affinité pour le 7-céto-oxystérol, il est exprimé dans la rétine.
Les oxystérols participent au développement de la chronicité de l’état inflammatoire par la production de cytokines (IL1β ; TNFα) et par la sur-expression des molécules d’adhésion VCAM-1 ; ICAM-1, E-Selectine.
a.3. Analogie des mécanismes pro-inflammatoires :
Dans l’athérosclérose et la DMLA, il semble qu’il existe une corrélation positive entre les taux de la protéine C-réactive (CRP) et certains marqueurs de l’inflammation tels que IL-6. D’autre part, le gène codant pour les iso-formes du facteur H du complément a été identifié comme un des facteurs génétiques qui prédispose aux lésions de la DMLA.
Ce facteur H module la réponse inflammatoire. L’inflammation chronique locale est stimulée par les dépôts de substance extracellulaire, elle est impliquée dans les pathologies dégénératives du vieillissement : Alzheimer, DMLA, athérosclérose.
Les protéines du complément sont des facteurs chémo-attractifs pour les macrophages identifiés dans les drusen.
Les MCP-1 (monocyte-chemotatic-protéine) sont impliqués dans la DMLA et l’athérosclérose en favorisant la production de cytokines pro-angiogéniques dans les cellules vasculaires.
Au cours de la formation de la plaque d’athérome, les protéines d’adhésion VCAM-1 et ICAM-1 sont exprimées à la surface de l’endothélium vasculaire. Ces protéines d’adhésion se lient aux récepteurs des cellules inflammatoires (monocytes), elles facilitent leur adhésion et leur pénétration dans l’intima artérielle. Les cellules endothéliales des capillaires choroïdiens expriment également des ICAM-1 et participent ainsi à l’inflammation chronique du RPE.
Références bibliographiques sur demande à Cette adresse e-mail est protégée contre les robots spammeurs. Vous devez activer le JavaScript pour la visualiser.
b. Polymorphisme génétique :
b.1. Polymorphisme des gènes impliqués dans le métabolisme des liquides et dans l’inflammation :
L’athérosclérose et la DMLA sont des maladies multifactorielles et polygéniques. Des étude récentes ont montré l’existence de polymorphismes génétiques et d’expression phénotypique pour lesquelles les corrélations sont difficiles à établir pour l’athérosclérose et la DMLA car parfois considérées comme contradictoires notamment en ce qui concerne le métabolisme des lipides.
Dans la DMLA, quatre facteurs de prédisposition génétique sont connus : le gène de l’APOE, le gène ABCA4, le gène CFH, et le gène PLEKHA-1.
Le gène de l’APOE et le gène CFH peuvent intervenir dans l’étiogenèse de ces deux pathologies dégénératives liées à un état inflammatoire chronique.
b.2. APOE et athérosclérose :
L’APOE est une des protéines constitutives des lipoprotéines HDL, LDL et VLDL impliquées dans le métabolisme des lipides. L’APOE est un ligand du récepteur de LDL.
Le gène de l’APOE localisé sur le chromosome 19, est polymorphe, 3 allèles déterminent 3 isoformes : APOE2, APOE3, APOE4 avec 6 phénotypes/génotypes.
Les isoformes se distinguent par la substitution d’un seul acide aminé (cystéine/arginine) ce qui modifie leur comportement biochimique vis à vis du cholestérol. Ainsi, l’isoforme APOE4 a une affinité plus élevée au récepteur des LDL, il est associé à une augmentation du cholestérol dans les LDL et à une élévation du risque cardiovasculaire. Les sujets porteurs de l’allèle ε-4 ont une clairance plus rapide des LDL plasmatiques.
L’isoforme ε-2 est associée à une diminution du cholestérol liée aux LDL et à une réduction du risque d’athérome.
L’isoforme ε-4 est également associé aux maladies neurodégénératives, telles qu’Alzheimer.
L’allèle APO ε-4 serait un marqueur indépendant prédictif du risque coronarien chez les sujets asymptomatiques et il est impliqué à 91% dans le risque de maladie d’Alzheimer.
En Europe, la fréquence de l’allèle ε-4 augmente du sud vers le nord et suit le gradient de fréquence des maladies cardiovasculaires. L’effet de polymorphisme de l’APOE dans les pathologies dégénératives cardiovasculaires et neurologiques suggère que le fait de ne pas posséder l’allèle ε-4 pourrait être un facteur de longévité, en effet, la fréquence de cet allèle est plus rare chez les centenaires. Cet allèle ε-4 correspondrait à l’allèle ancestral et que dans certaines circonstances, cet allèle extrêmement conservé pourrait avoir eu ou aurait encore des effets favorables qui expliqueraient sa persistance au fil des générations. Les travaux de E. Souied, de C. Klaver ont démontré son effet protecteur dans la DMLA oxydative.
Ce polymorphisme induit des effets opposés : augmentation du risque d’athérosclérose et diminution du risque de la DMLA. Ces effets opposés observés pour les différents isoformes des APOE peuvent s’expliquer par des propriétés de fixation et d’internalisation des LDL et HDL qui seraient « cellules spécifiques » dans les cellules de l’épithélium pigmentaire rétinien, des neurones et des astrocytes du système nerveux central.
b.3. APOE et DMLA :
L’APOE a été mise en évidence dans les drusen. Souied et col, Klaver et col ont démontré l’association du gène codant pour l’APOE à la DMLA. De plus, les porteurs de l’allèle ε-4 caractéristique de l’isoforme APOE ε-4 sont protégés de l’apparition de la DMLA avec des odds ratio allant de 0,34 à 0,43.
Cette protection concerne la DMLA exsudative caractérisée par les drusen séreux. Un individu porteur de l’allèle ε-4 a un risque relatif d’être atteint de drusen séreux avec DMLA exsudative 4 à 5 fois moindre qu’un sujet non porteur de cet allèle ε-4.
La composition chimique au niveau des acides aminés de l’APO ε-4 confère des propriétés biochimiques particulières qui rendent l’APOE 4 incapable de former des dimères de grosses particules lipidiques, ce qui faciliterait une meilleure clairance du cholestérol au travers de la membrane de Bruch. De plus la présence des deux charges électriques positives peuvent expliquer une meilleure élimination des débris cellulaires au travers la membrane de Bruch qui est devenue hydrophobe avec le vieillissement.
E. Loane et al. ont montré une corrélation entre le génotype APOE ε-4 et le MPOD (Macular Optical Density). Cette association peut expliquer le rôle protecteur de ce profil d’apolipoprotéine et le transport, la capture de la Lutéine et de la zéaxanthine par la rétine.
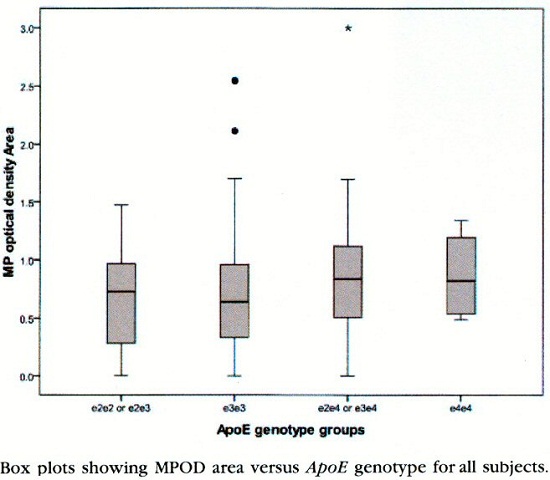 Box plots showing MPOD area versus ApoE genotype
Box plots showing MPOD area versus ApoE genotype
Selon Klaver et al., le polymorphisme APOE ε-4 présente un risque faible de DMLA , les sujets porteurs du génotype APOE ε-2 ont un risque élevé.
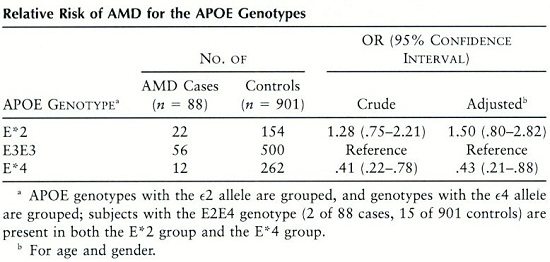 Relative risk of AMD for the ApoE genotypes
Relative risk of AMD for the ApoE genotypes
b.4. Facteur H du complément (CFH) athérosclérose et DMLA :
Certaines études récentes ont corrélé l’athérosclérose à l’activation du Complément. Le facteur H du complément CFH est une protéine qui régule la voie alterne du Complément, il assure une protection des tissus contre une activation inadaptée du système Complément.
Dans l’athérosclérose et la DMLA, on a observé un polymorphisme du gène CFH codant pour le facteur H ; il se caractérise par une substitution de la tyrosine par une histidine en position 402 (Y402H). Cette modification dans la structure de la protéine CFH induit une diminution de l’activité inhibitrice de la voie alterne du Complément.
En effet, ce variant en position 402 se situe dans le site de la liaison à l’héparine, à la protéine C réactive (CRP) et à la protéine M ; la substitution de la tyrosine (acide aminé neutre) par l’histidine chargée positivement peut altérer cette liaison à la CRP. La colocalisation de CFH, CRP, et de protéoglycanes dans la couche superficielle de l’intima artérielle suggère que CFH protégerait d’une activation excessive du Complément lors d’état inflammatoire. La présence du polymorphisme Y402H peut donc diminuer cette action protectrice et favoriser les lésions vasculaires et la néovascularisation observées dans la forme humide de la DMLA et l’athérosclérose suite à l’action pro-inflammatoire et pro-angiogénique des VCAM-1,ICAM-1, intégrines LFA1 et VLA-4 exprimées à la surface de l’endothélium vasculaire.
Il est actuellement reconnu qu’une élévation modérée et chronique de la CRP représente un facteur de risque de maladie cardiovasculaire ; la CRP n’est pas seulement le témoin d’une « micro-inflammmation » dans la plaque d’athérome, mais elle constitue un des acteurs de l’athérogénèse. La liaison aux groupements phosphorylcholines présents sur les LDLox, facilite la capture des LDL par les macrophages et permet l’expression des molécules d’adhésion dans les cellules endothéliales.
b.5. Gène SCARB-1 : transport des lipides et de la Lutéine :
SCARB-1 est impliqué dans le métabolisme de trois molécules clefs en relation avec l’étiologie de la DMLA et de l’athérome : le cholestérol, la lutéine et la vitamine E.
Le gène SCARB-1 code pour le récepteur de surface multiligand SRB-1 connu pour la capture sélective et l’efflux du cholestérol, de la vitamine E et de la Lutéine.
Ce gène SCARB-1 joue un rôle ubiquitaire, il participe au métabolisme de ces trois molécules lipophiles depuis leur capture par les entérocytes jusqu’au passage dans la rétine.
Le variant rs 5888 du gène SCARB-1 serait associé à une augmentation du risque de DMLA, et de maladies cardiovasculaires.
A. During et al. ont bien montré le passage préférentiel de la Lutéine dans les cellules rétiniennes via un mécanisme SCARB-I dépendant, lié au transport des HDL.
S. Acton et al. ont identifié d’autres variants du gène SRB-1 impliqués dans les dyslipidémies et l’obésité en particulier chez la femme en pré-ménopause. Les analyses de ces auteurs montrent un polymorphisme du gène SRB-1 localisé en 12q24 dans les populations de race blanche, des corrélations peuvent être établies entre différents SNP (Single Nucleotide Polymorphism), les lipides plasmatiques et des mesures anthropométriques, l’IMC,….
Ces SNP sont sexe-spécifiques, chez l’homme, l’allèle 2 est associé à un profil lipidique moins athérogène ; chez la femme pré-ménopausée, le polymorphisme de l’intron 5 est corrélé à l’IMC.
Ainsi S. Acton et al. ont montré que la présence de cet haplotype 121 augmentait l’IMC des femmes de 2.2 kg/m2, soit 6 kg en poids corporel. Cette hausse de l’IMC résulte en une augmentation du risque cardiovasculaire de 17,6%, avec une incidence importante pour le diabète de type II. Ces SNP sont corrélés au taux de cholestérol des HDL et des LDL, et une surexpression hépatique du SRB-1 induit une excrétion accrue du cholestérol dans la bile et réduit l’absorption du cholestérol alimentaire. D’autre part, l’ARNm du SRB-1 est exprimé dans le tissus adipeux dans lequel le SRB-1 régule l’entrée des lipides dans les adipocytes. De plus, l’expression du SRB-1 dépend des glucocorticoïdes et des œstrogènes ; ces hormones jouent de ce fait un rôle important dans la genèse de l’intolérance au glucose, des dyslipidémies et de l’augmentation de l’IMC.
F. Rodriguez et al. ont également observé des polymorphismes des gènes du SRB-1, de l’enzyme paraoxonase (PON), du CD36 et du CLA-1 (lysosomal integral membrane proteine II analogous -1). Ces protéines jouent un rôle central dans la capture du cholestérol et des esters de cholestérol oxydés dans les HDL.
Les propriétés antioxydantes des HDL ont été attribuées à l’activité enzymatique de la paraoxonase 1 (PON1) pour lesquelles il existe des variants alléliques ainsi l’effet protecteur anti-athérogène des HDL est moindre chez les porteurs du génotype PON RR par rapport aux porteurs du génotype PON1-QQ.
P. Borel et al. ont identifié des variants des gènes BCMO1 et CD36 qui étaient associés aux teneurs plasmatiques en Lutéine et dans la rétine.
Références bibliographiques sur demande à Cette adresse e-mail est protégée contre les robots spammeurs. Vous devez activer le JavaScript pour la visualiser.
c. Tropisme spécifique de la lutéine pour les cellules vasculaires et l’épithélium pigmentaire rétinien :
c.1. Spécificité et structure moléculaire de la Lutéine :
Les propriétés protectrices de la Lutéine et de la zéaxanthine ont été largement démontrées dans différentes études relatives à la protection cardiovasculaire et à la DMLA : Los Angeles, Atherosclerosis Study (cf page 1 et 2), Rotterdam Study, CARDIA Study (cf page 1 et 2), The Aric Study (cf page 1 et 2), Monica Toulouse Belfast (cf page 1 et 2), The Veterans Last Study (cf page 1 et 2), The Luxea Study, etc,…
Ces propriétés protectrices de pathologies dégénératives sont dues au pouvoir antioxydant de la Lutéine et de la zéaxanthine.
La Lutéine et la zéaxanthine sont des oxy-caroténoïdes, l’hydroxylation au bout de la chaine isoprénique procure un pouvoir anti-radicalaire plus puissant à ces deux caroténoïdes par rapport au β-carotène et lycopène.
Cette configuration « hydroxylée » permet à la Lutéine une pénétration en profondeur dans la phase hydrophobe des lipides membranaires ou des lipoprotéines HDL et LDL.
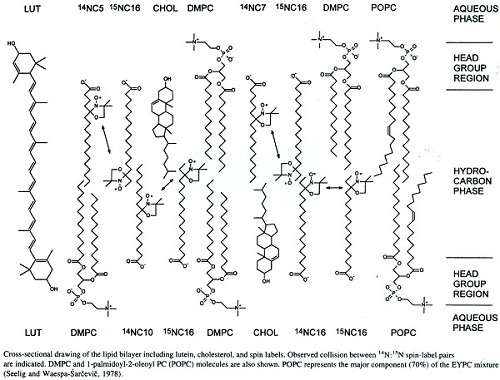 Effects of Lutein and Cholesterol on Alkyl Chain Bending in Lipid Bilayers
Effects of Lutein and Cholesterol on Alkyl Chain Bending in Lipid Bilayers
La Lutéine et le Zéaxanthine se retrouvent en abondance dans les membranes riches en cholestérol et acides gras polyinsaturés (DHA et EPA) des épithéllias vasculaires et rétiniens ou encore dans les lipoprotéines. Le cholestérol et la Lutéine jouent des rôles similaires dans la fluidité membranaire.
La polarité cellulaire de la Lutéine est « membrane spécifique ». Elle se concentre dans les membranes les plus épaisses et qui nécessitent une grande fluidité. Cette spécificité de la Lutéine réside dans sa configuration spatiale au sein des membranes.
Ainsi le cholestérol par sa plus petite taille, n’est présent que dans un seul feuillet lipidique de la bicouche, tandis que la Lutéine par la présence des radicaux hydroxyles polaires et par la longueur de sa chaine isoprénique adopte une position transmembranaire et constitue ainsi un « rivet » qui maintient en bon ordre les chaines alkyles des lipides.
Le cholestérol crée des poches dans la partie centrale des chaines lipidiques qui sont facilement occupées par l’O2. A l’opposé, la Lutéine agit comme une barre d’ancrage et interagit avec les deux feuillets lipidiques diminuant ainsi le transport d’O2 au plus profond de la membrane lipidique.
La Lutéine réduit l’isomérisation trans-gauche en renforçant la configuration trans dans la chaine alkyle sur toute sa longueur ce qui limite le transport d’O2 et la peroxydation lipidique.
Wisnieska et al. ont démontré que cette orientation perpendiculaire de la Lutéine affectait significativement les propriétés physiques des membranes, incluant leur fluidité, l’ordre des chaines alkyles, leur rotation, l’hydrophobicité de la membrane intérieure et la vitesse de transport de l’O2 dans la membrane.
Ces modifications des propriétés physiques de la membrane sont considérées comme essentielles dans le mode d’action protecteur de la peroxydation lipidique par la Lutéine et la zéaxanthine dans l’épithélium vasculaire et rétinien et dans les HDL et LDL.
La Lutéine agit en règle générale comme scavanger non-stœchiométrique par effet de « quenching » de l’oxygène singulet.
c.2. Propriétés anti-inflammatoires de la Lutéine :
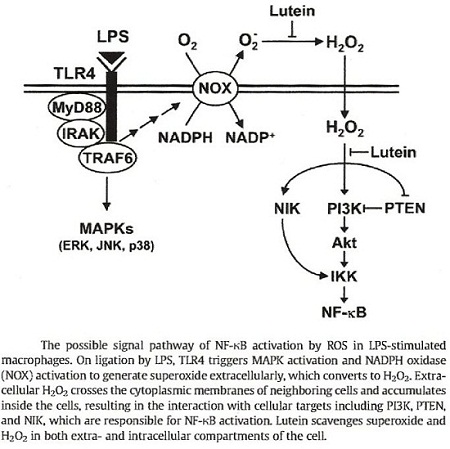
c.2.2-1 Action inhibitrice du NF-k-B :
Le facteur de la transcription nucléaire NF-k-B régule l’expression des gènes impliqués dans l’inflammation et la prolifération cellulaire.
Dans les cellules quiescentes, le NF-k-B se trouve sous forme inactive dans le cytosol, car il est lié à un inhibiteur : l’IkB. Lors d’une modification du potentiel redox dans le cytosol, par un excès de radicaux libres : oxygène singulet, H2O2 ou par la présence d’endotoxines, de LPS (Lipopolysaccharides bactériens), des kinases : l’I-k-β kinase (IKK), la MAP kinase, la JNK induisent la phosphorylation et la dégradation de I-k-β libérant ainsi le NF-k-B.
Le NF-k-B active la transcription nucléaire des gènes du TNF-α de l’IL-1β, de l’INOS, de la COX2 et des promoteurs de l’inflammation des molécules d’adhésion ICAM, VCAM.
Tous ces médiateurs seront responsables de l’inflammation chronique et de la néovascularisation observées dans les pathologies dégénératives.
Ji-Hee Kim et col. ont mis en évidence dans des macrophages stimulés par le LPS, l’inhibition du système kinase activateur du NF-k-B.
La Lutéine diminue la teneur intracellulaire en oxygène singulet et en H2O2 régule le potentiel redox et bloque de ce fait l’expression des gènes responsables de sérine /thréonine phosphatase contenant des radicaux thiols « redox-sensibles »
 Effects of Lutein and Cholesterol on Alkyl Chain Bending in Lipid Bilayers
Effects of Lutein and Cholesterol on Alkyl Chain Bending in Lipid Bilayers
Dans une autre étude, Xue-Hai Jin et col. avaient montré l’action anti-inflammatoire de la lutéine dans l’uvéite induite par endotoxines. Selon ces auteurs, la Lutéine bloque la dégradation de l’I-k-β dans le cytosol des macrophages activés. La lutéine pourrait aussi inhiber d’autres facteurs de transcription induits par le LPS et supprimer l’expression de médiateurs pro-inflammatoires rencontrés dans l’uvéite tels que les iNOS-NO, COX2 –PGE2.
K. Izumi-Nagai et col. ont identifié les mécanismes d’action anti-inflammatoire de la Lutéine dans la pathogénèse de la néovascularisation choroïdale rencontrée dans la DMLA.
Cette étude a montré les mécanismes cellulaires et moléculaires de la suppression de la néovascularisation choroidale par le traitement avec la Lutéine.
La Lutéine conduit à une suppression significative in vivo et in vitro de l’infiltration des macrophages et des molécules pro-angiogéniques telles que : ICAM-1, MCP-1 et VEGF.
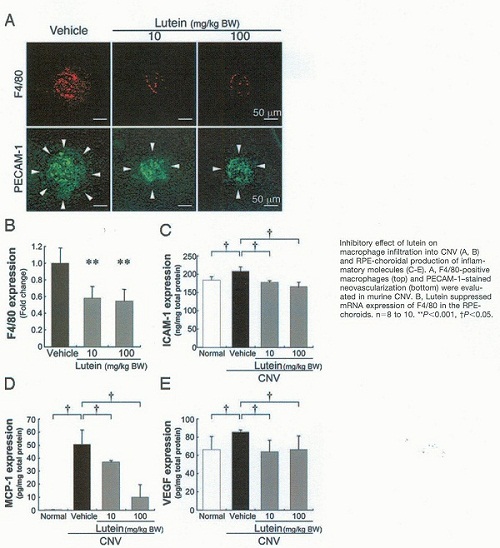 Inhibitory effect of lutein on macrophage infiltration
Inhibitory effect of lutein on macrophage infiltration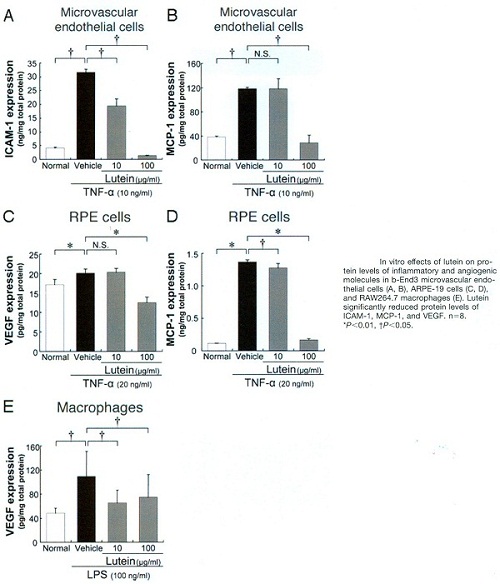 Macular Pigment Lutein is Antiinflammatory in Preventing Choroidal Neovascularization 1
Macular Pigment Lutein is Antiinflammatory in Preventing Choroidal Neovascularization 1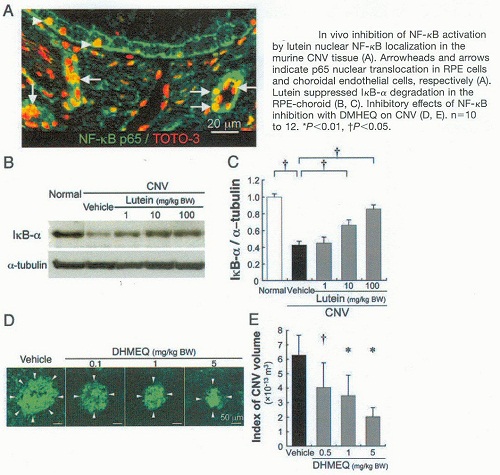 Macular Pigment Lutein is Antiinflammatory in Preventing Choroidal Neovascularization 2
Macular Pigment Lutein is Antiinflammatory in Preventing Choroidal Neovascularization 2K.Izumi-Nagai, et al. Macular Pigment Lutein is Antiinflammatory in Preventing Choroidal Neovascularization. Arterioscler Thromb Vasc Biol. 2007;27:2555-2562
c.2.2-2 Action inhibitrice du peroxinitrite :
Le peroxynitrite est un puissant oxydant formé in vivo à partir du NO et de l’O2.
Il réagit avec les lipides et les acides aminés détruisant les complexes protéino-lipidiques des lipoprotéines plasmatiques. Les caroténoïdes (β-carotène, lycopène) et les oxycaroténoïdes (Lutéine et zéaxanthine) en particulier sont des scavangers du peroxynitrite.
O. Panasenko et col. ont démontré dans une étude in vitro l’efficacité de la Lutéine qui est comparable à celle des biothiols (glutathion et cystéine) reconnus comme les scavangers physiologiques du peroxynitrite.
Les caroténoïdes et oxycaroténoïdes protègent les lipoprotéines plasmatiques des modifications de leur structure par les nitrosylations du peroxynitrite et les exposent de ce fait moins au récepteur scavanger des macrophages et donc évitent la formation des cellules responsables des lésions athéromateuses précoces.
Références bibliographiques sur demande à Cette adresse e-mail est protégée contre les robots spammeurs. Vous devez activer le JavaScript pour la visualiser.
d. Etudes cliniques récentes : la lutéine efficace à quelle dose ? :
S. Richter et col. ont confirmé dans « the Zeaxanthine and Visual Study » (ZVF) FDA IND que la dose de 9 mg libres de Lutéine était bien adaptée pour améliorer les paramètres visuels de faible contraste et d’éblouissement lumineux observés dans la DMLA.
Par rapport aux doses de 9 mg libres de Lutéine associées à 8 mg de zéaxanthine, la supplémentation à la dose de 9 mg libres de Lutéine apparaissait la plus performante. Les résultats de l’étude ZVF confirment ceux obtenus dans l’étude LAST avec une supplémentation dosée à 10 mg libres (20 mg esters) de Lutéine (*)
D’autres études récentes ont confirmé l’intérêt des supplémentations de 10 mg libres (20 mg esters) de Lutéine pour relever de façon significative et durable la densité pigmentaire de la macula. The Luna Study (**); The Lisa Study.
Dans l’étude LISA, on a pu établir une corrélation entre l’augmentation pigmentaire de la macula et une amélioration de l’acuité visuelle.
Dans une étude randomisée réalisée pendant 2 ans sur 145 patients atteints de DMLA, S. Piermarocchi et col. ont constaté que les patients traités par 10 mg libres (20 mg esters) de Lutéine associées à 1 mg de zéaxanthine et 4 mg d’astaxanthine présentaient une meilleure stabilisation de l’acuité visuelle, de la sensibilité de contraste et de la fonction visuelle par rapport aux patients non supplémentés. (***)
Selon Th. Desmettre et J.M. Lecerf, les doses journalières préconisées actuellement sont de 10 mg libres (20 mg esters) de Lutéine et 2 mg de zéaxanthine. La revue des études cliniques faites par S. Carpentier montre également l’intérêt des doses supérieures ou égales à 10mg d’oxycaroténoïdes.
(*) S.P. Richer et al. Randomized, double-blind, placebo-controlled study of zeaxanthin and virtual function in patients with atrophic age-related macular degeneration. Optometry 2011,82:667-680(**) M. Zeimer et al. The macular pigment : shoft and intermediate term changes of macular pigment optical density following supplementation with lutein and zeaxanthine and co-antioxydants. The Luna study. Ophtalmologue. 2009. Jan; 106 (1):29-36.
(***) S. Piermarocchi et al. Carotenoids in Age-related Maculopathy Italian study (CARMIS): two-year results of a randomized study. Eur. J. Ophtalmol. 2011.Oct. 17: 0
____________
D’autres nutriments tels que les acides gras oméga-3 à longues chaines (DHA - EPA) l’α-tocophérol, le zinc et le sélénium, présentent également un intérêt dans la régulation physiologique des maladies dégénératives.
- Antioxydant et : stress oxydant
- Antioxydant et : vitamines antioxydantes
- Antioxydant : la Grenade
- Antioxydant : le Coenzyme Q10
- Antioxydant : le Glutathion
Références bibliographiques sur demande à Cette adresse e-mail est protégée contre les robots spammeurs. Vous devez activer le JavaScript pour la visualiser.